|
摘 要
动物肠道内栖息着大量微生物,肠道微生物及其代谢产物积极参与肠道免疫稳态的建立,对防控肠道疾病具有积极影响,在动物健康中发挥不可或缺的调节作用。本文对肠道微生物及其代谢产物对宿主肠道免疫的影响进行综述,旨在为调控肠道免疫、治疗动物疾病及保障机体健康提供思路和参考。
1 肠道微生物及代谢产物
1.1 肠道微生物的分布
在已鉴定的肠道菌群中,拟杆菌门和厚壁菌门的数量占肠道菌群总量90%以上,其中专性厌氧菌数量约是需氧菌数量的1 000倍。由于不同肠段内环境不一样,肠腔中微生物数量与组成均有差异。十二指肠中的微生物数量最少,主要原因是胃酸与胆汁排出后首先进入其中,导致内环境pH较低,且过多的肠液和肠段快速蠕动也不利于细菌定植。与十二指肠相比,空肠和回肠中的肠液更少,肠段蠕动速度减缓,氧化还原电位降低,促进厌氧菌定植,故空肠和回肠内的肠道微生物数量有所增加。盲肠具有远端闭塞不通、肠段蠕动速度缓慢和pH 呈弱碱性等特点,是细菌定植的主要场所,其中优势菌群为双歧杆菌(Bifidobacterium)、肠杆菌(Enterobacteriaceae)和肠球菌(Enterococcus)。
1.2 肠道微生物产生的代谢产物
动物肠道无法直接消化吸收的饮食成分,经肠道微生物作用后,转化为宿主代谢中的功能性补充物质。微生物代谢产物分为3 类:第一类经肠道微生物直接对食物成分进行消化或发酵产生,如SCFAs。SCFAs 由小肠厌氧微生物发酵结构性碳水化合物生成,具有促进肠道黏液分泌、提供结肠肌细胞能量等作用。第二类由宿主产生后经肠道微生物修饰后的代谢产物,如次级胆汁酸。次级胆汁酸由初级胆汁酸经肠内7-α/β 脱羟基细菌解离、转化而来,有助于动物构建肠道环境稳态。第三类为肠道微生物自生合成的代谢产物,如PSA。PSA 主要由BF产生,是一种荚膜多糖,通过激活Toll 样受体2(TLR2)信号通路诱导Treg 细胞产生具有抗炎作用的IL-10,抑制胃肠道炎症的发生。综上所述,微生物可以通过其代谢产物调节宿主胃肠道免疫系统的正常生理功能(表1)。

2 肠道微生物对免疫细胞和免疫球蛋白的影响
2.1 肠道微生物对分泌型球蛋白A(sIgA)分泌的影响肠道微生物通过依赖T细胞和非依赖T细胞2种方式调控sIgA的产生(图1)。依赖T细胞的sIgA产生主要发生在派尔集合淋巴结中,其通过B 细胞与负载抗原的DCs 相互作用后产生sIgA。捕获抗原的DCs刺激CD4+T 细胞增殖分化为滤泡辅助性T(Tfh)细胞,Tfh 细胞主要以CD40 配体和IL-21作为信号,诱使B细胞表达胞嘧啶核苷脱氨酶(AID),促进B细胞分化为IgA+B细胞,进而分泌大量sIgA。非依赖T细胞的sIgA产生主要在肠黏膜固有层和孤立淋巴滤泡中。肠道菌群可通过触发肠上皮细胞中TLRs 直接刺激DCs和滤泡树突状细胞产生增殖诱导配体(APRLI)和B细胞的活化因子(BAFF),诱导B 细胞中的AID表达和IgA类抗体转换,提高B细胞转化为IgA+B细胞的速率,显著增强sIgA合成能力。

肠黏膜固有层中IgA+浆细胞产生二聚体IgA(dIgA),dIgA 相互聚合形成多聚体IgA(pIgA),随后与多聚免疫球蛋白受体(pIgR)结合转运至肠上皮表面顶端,之后dIgA与pIgR中的分泌成分从结合物中分离,形成sIgA。sIgA接触肠腔内微生物、膳食成分等物质后,在这些物质表面形成一层外膜,抑制肠腔致病菌直接接触肠壁,使其无法黏附肠上皮细胞失去定植能力,从而维持机体肠道屏障的完整性,抵御其他病原微生物入侵。
2.2 肠道微生物对T细胞的影响
肠腔内特定的共生细菌可以促进T细胞不同亚群的生成。抗原刺激使固有层中CD8+T细胞向CD4+T细胞增殖分化后,CD4+T细胞又可特异性增殖分化为不同亚群,如Treg细胞、Tfh细胞和Th17细胞等。其中,Treg细胞的增殖分化可以减轻因机体对致病菌的过度免疫反应而引起的黏膜损伤,如BF诱导Treg细胞增殖分化产生IL-10,保护肠道免受肝螺杆菌(Helicobacter hepaticus )入侵感染。Th17细胞具有介导炎症反应的作用,其与Treg 细胞在功能上具有相互制约的作用。近年来大量研究表明,通过肠道微生物诱导产生的Treg细胞与Th17细胞可以维持肠道在免疫耐受与炎症反应之间的平衡,有利于建立肠黏膜免疫稳态。
2.2.1 肠道微生物对Treg细胞增殖分化的影响
Treg细胞来源于胸腺和外周免疫器官,其增殖分化主要受CD25和叉头状螺旋转录因子3(FoxP3)分子调控。肠道Treg 细胞存在于肠黏膜和淋巴组织中,抑制T细胞过度反应,调节自身免疫。外周Treg细胞(pTreg)聚集于结肠,常见表达形式为RORγt。无菌小鼠体内缺乏RORγt+ pTreg 细胞,说明微生物可以诱导产生pTreg 细胞。Atarashi等将正常人体17种梭菌混合,定植于无菌小鼠体内,结果显示pTreg细胞水平显著升高,促进IL-10 和细胞毒T淋巴细胞相关抗原4的合成释放,共同抵御小鼠结肠炎。研究表明,BF可以分泌外膜囊泡,通过非典型自噬途径向免疫细胞传递免疫调节分子,诱导产生IL-10,表达FoxP3+细胞,促使CD4+T 细胞向Treg 细胞增殖分化,抑制结肠炎。
2.2.2 肠道微生物对Th17 细胞增殖分化的影响Th17细胞的应答时间早于其他T 细胞,且其可以在肠道内积聚,这表明Th17细胞的发育由肠内机制调节。与此相契合的是,使用抗生素处理后的小鼠或者无菌小鼠,肠内Th17细胞数量均急剧下降。因此,肠道微生物在调控Th17 细胞的增殖分化中具有重要作用。研究表明,肠道中SFB 具有钩状结构,钩尖黏附小鼠肠上皮细胞形成内吞囊泡,可快速将SFB 的菌体蛋白从肠上皮细胞的顶端运送至基部,直接促使血清淀粉样A 蛋白(SAA)产生,进而使黏膜固有层中DCs产生IL-6和IL-23,诱导Th17 细胞增殖分化。同时,Mao等证明SFB 定植肠道后产生IL-23,刺激固有淋巴细胞(ILCs)与Th17细胞共同分泌IL-22,间接上调SAA1和SAA2基因表达,并激活MAPK通路释放抗菌肽和β防御素,保证肠道屏障结构与功能的完整性,阻碍病原微生物入侵机体。除SFB外,肠内白色念珠菌(Candida albicans)、鼠柠檬酸杆菌(Citrobacter ratus)及肠出血性大肠杆菌(Escherichiacoli O157: H7)等致病菌均可黏附肠上皮促使Th17 细胞分化。由此看来,肠道微生物可以通过对肠上皮细胞的黏附作用诱导T 细胞向Th17 细胞增殖分化,对机体炎症迅速做出反应,加强宿主对病原微生物的防御作用(图2)。

3 肠道微生物代谢产物与肠黏膜免疫
3.1 SCFAs 与肠黏膜免疫
SCFAs包括乙酸、丙酸、丁酸等,SCFAs不仅是微生物本身的主要能量来源,还为肠上皮细胞提供能量。此外,SCFAs还能抑制组蛋白去乙酰酶(HDACs)活性,促使肠上皮细胞紧密连接,参与调节机体免疫功能。SCFAs通过抑制HDACs 活性影响Treg 细胞发育。HDAC9的抑制作用使FoxP3表达增加,促使Treg 细胞增殖分化,增强Treg 细胞介导的结肠炎抑制作用。研究表明,SCFAs 与游离脂肪酸受体2(FFAR2)特异性结合后减弱肠道中性粒细胞的趋化作用,降低脂多糖刺激下中性粒细胞产生的促炎细胞因子水平。Zhou 等研究发现脂多糖刺激巨噬细胞后,丁酸能限制促炎细胞因子在结肠固有层中合成与分泌,降低肠道炎症的发生几率,同时结肠隐窝存在丁酸代谢,防止高浓度丁酸盐对隐窝底部干细胞产生毒害效应,维护动物肠黏膜免疫细胞的更新能力。
SCFAs 通过完善肠上皮细胞屏障功能维持黏膜免疫。Wrzosek等研究发现,无菌小鼠接种产生SCFAs 的泰氏杆菌(Bacteroides thetaiotaomicron) 和粪便杆菌(Faecalibacterium),能有效诱导杯状细胞分化,促进黏液产生,有效阻止致病菌定植肠道,保障肠道健康。SCFAs还能稳定肠上皮细胞连接稳定,维护肠道屏障完整。定植长双歧杆菌(Bifidobacteriumlongum)产生高水平乙酸,增强肠上皮细胞间相互连接,提高肠道对病原体的防御能力。
3.2 次级胆汁酸与肠黏膜免疫
胆汁酸由肝脏合成储存于胆囊,动物采食后胆囊收缩,胆汁酸被释放进入十二指肠,称为初级胆汁酸。肠道腔内乳酸杆菌、双歧杆菌等利用胆盐水解酶对初级胆汁酸进行早期解离后,再与具bai 基因的梭菌发生7-α脱羟基反应生成次级胆汁酸,包括石胆酸(LCA)、脱氧胆酸(DCA)和熊脱氧胆酸(UDCA)等。研究证明,患炎症性肠病(IBD)的小鼠体内厚壁菌门数量减少,阻碍胆汁酸早期解离、转化及脱硫过程,显著降低次级胆汁酸转化率,导致3-OH硫化胆汁酸积聚肠内,加重炎症反应。
Den Bossche等将UDCA 灌胃于葡聚糖硫酸钠诱导IBD 小鼠后,使肠内厚壁菌门与拟杆菌门比例恢复正常,并有效降低小鼠肠道促炎细胞因子(IL-1β 、IL-6)水平,加速溃疡化上皮愈合,表明次级胆汁酸具备一定免疫调节作用。此外,研究表明,LCA 与DCA有效抑制80%肠道白色念珠菌菌丝、芽管及生物膜的形成,阻止其生长繁殖,维持动物肠道健康。Jason 等试验证明,LCA和DCA可增强肠内7-α 脱羟基细菌所分泌抗生素的活性,解除外源抗生素治疗艰难梭菌后产生的环状二肽抑制其他菌群生长,阻止艰难梭菌异常增殖所导致的肠黏膜免疫极化现象出现。同时,次级胆汁酸作为胆汁酸家族一员,也具备激活法尼醇X受体及G 蛋白偶联胆汁酸受体5通路的功能,发挥维持肠道稳态及黏膜屏障完整性的作用。综上所述,适宜浓度的次级胆汁酸对机体肠黏膜免疫建立具有重要意义。
3.3 PSA 与肠黏膜免疫
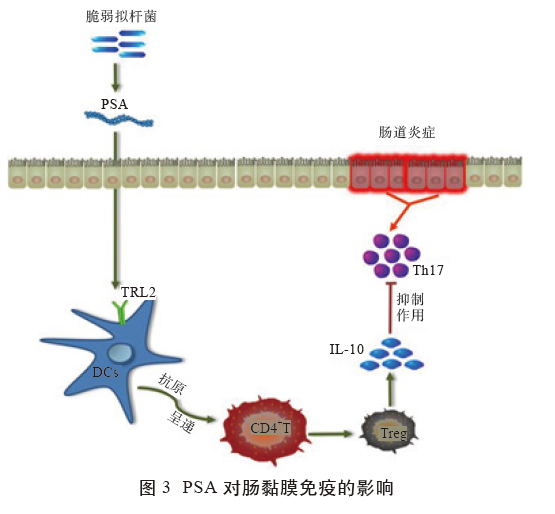
PSA为囊状两性离子多糖,主要由BF产生,常见于结肠黏液层,通过激活DCs和Treg 细胞合成分泌抗炎细胞因子,调控宿主免疫系统功能。一方面,PSA 依赖肠壁通透性进入固有层,被DCs内化吸收后将抗原呈递至CD4+T细胞,导致其分泌IL-10,诱使CD45+细胞释放IL-17A,抑制肠道T 细胞活性,尤其是Th17细胞,继而阻止炎症反应发生,保护罹患结肠炎小鼠的生理健康。另一方面,DCs 细胞表面的TLR2 也能被PSA 激活,促使T 细胞释放IL-10。此外,PSA还能刺激CD4+T细胞向FoxP3CD4+ T细胞或FoxP3-CD4+ Tr1 细胞方向分化,与细胞表面TLR2 特异性结合,促进IL-10的合成,缓解结肠炎。综上所述,PSA 通过直、间接方式作用于CD4+T细胞,促进IL-10分泌,抑制Th17细胞过度免疫应答,辅助动物抵御肠道疾病(图3)。
4 结 语
综上所述,肠道微生物定植后调节sIgA分泌及T细胞增殖,完善动物肠道免疫功能,同时产生大量代谢产物(SCFAs、次级胆汁酸和PSA等),在维持肠道稳态、辅助动物抵御疾病方面具有重要作用。肠道微生物及其代谢产物对动物肠道免疫的影响已被大量研究和阐释,但菌群种类及定植过程复杂,现有研究大多为孤立、单一角度的验证,仍不足以解释菌群与肠道免疫间复杂的动态关系。因此,肠道微生物及其代谢产物对宿主肠道免疫系统作用机制有待全面深入探索,相关研究也将为丰富机体肠内外疾病的防治理论提供参考依据。(参考文献略)
|
