|
PRRS弱毒活疫苗保护效果对同源性要求有多高?
向金梅 1,2 #,汪志恒 3 #,陈文钦 2,赖庆光 4,王雯熙 2,万春云 1,4 *(1.长江大学,湖北?荆州?434200;2.湖北生物科技职业学院,湖北?武汉?430000;3.罗牛山畜牧股份有限公司,海南?海口 570000;4.浙江美保龙生物技术有限公司,浙江?金华?321000)猪繁殖与呼吸综合征(Porcine Reproductive Syndrome,PRRS)也称猪蓝耳病,是世界上对猪场影响最严重的猪病;猪繁殖与呼吸综合征病毒(Porcine Reproductive Syndrome Virus,PRRSV)几乎在所有区域猪场都有流行,也是目前我国猪场最顽固、带来危害和经济损失最大的病原之一。
从血清学角度来看,PRRSV的血清型主要有1型和2型,在我国主要流行的是2型。如果仅从血清中和能力的角度来理解,在我国流行的PRRSV主要毒株对疫苗的选择性不高,其刺激机体所产生的抗体大多数不是中和抗体,也不产生高滴度的中和抗体,且该病毒本身也不能完全依赖于中和抗体进行控制,导致疫苗在实际使用中体现出极强的选择性,尤其是弱毒活疫苗,对毒株匹配的选择性更强,这也与活苗和灭活苗的激发免疫的不同机制有关系。
另一方面,因为PRRSV具有极强的变异性,导致该病毒变异频繁、毒株多样,其变异演化速率是RNA病毒中最快的。每隔几年我国流行的PRRSV主要亚型就会发生变化,而次一级的PRRSV亚型变化更多,究其原因,除病毒本身不保真、无校正能力的复制特性之外,造成 PRRSV 变异与演化复杂的机制还涉及病毒准种和毒株重组,目前越来越多的证据表明活苗的使用在其中发挥了重要作用。研究者认为,不恰当的弱毒苗使用是加快病毒变异的重要原因,使猪蓝耳病流行更加复杂化。
目前我国猪场主要流行的PRRSV毒株和亚系情况见图1,显示PRRSV流行毒株具有复杂性和多变性。展示出来的仅是冰山一角,实际情况更加复杂,除了PRRSV多毒株共同感染之外,随时也可能出现新的亚型。
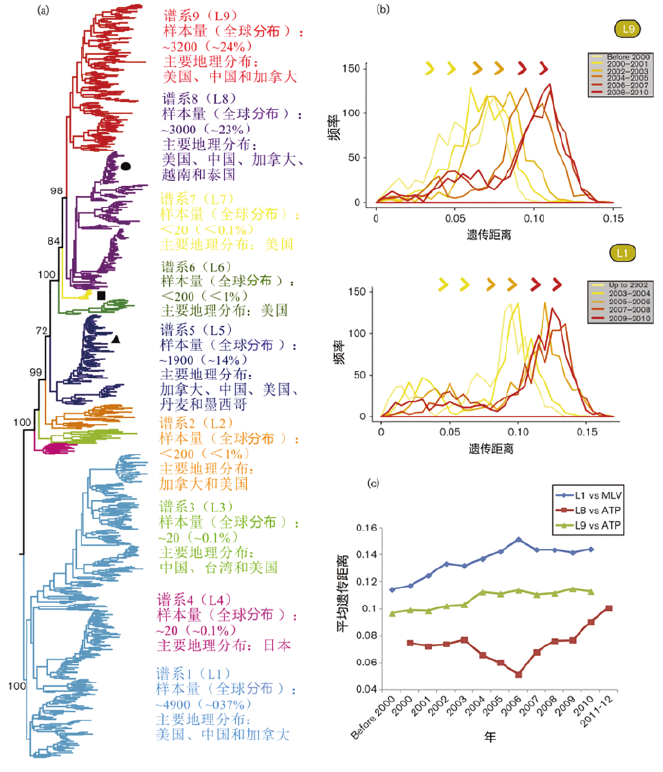
图1 基于ORF5分型的2型PRRSV主要谱系及随时间出现的变异和多样化趋势
PRRS弱毒活疫苗在世界上已经有较长的使用历史,在我国使用也有多年的历史。2006年后随着猪蓝耳病给我国猪场带来的损失越来越严重,2008年前后PRRS疫苗免疫逐渐被猪场接受,在这个时间段猪场使用过PRRS灭活疫苗、弱毒活疫苗,最终因为PRRS弱毒活疫苗效果更佳而被猪场认可和接受。PRRS的免疫一度被纳入国家动物疫病的强制性免疫范畴,PRRS弱毒活疫苗的使用也确实解决了国内猪场的猪蓝耳病防疫问题。
随着PRRS弱毒活苗的使用,以及弱毒活疫苗毒株范围的逐渐加大,近年来我国PRRS发病和流行的形势逐渐复杂化。我国使用过的PRRS弱毒活疫苗毒株主要有JXA/1R、R98、VR2332,此外还有天津株、湖南株、M2等等。PRRS弱毒活疫苗毒株多,使用范围广,占了猪蓝耳病疫苗的大半江山。目前,很多猪场已经非常难以控制,PRRS频繁出现波动,频繁出现新的毒株,使我国PRRS情况进入复杂循环的怪圈。
如前所述,PRRS弱毒活疫苗的保护效果对毒株同源性的要求很高,目前尚在商业推广使用的PRRS-MLV对基因同源性高的病毒具有更高的保护效力,这已经是PRRS控制的一个共识。蓝耳活苗的保护效力对同源性的要求,早在上个世纪九十年代就发现了,Collins等人(1992)评估了美洲型和欧洲型弱毒疫苗对来源不同的3株欧洲型田间毒入侵时免疫保护力的高低,证实若所攻的病毒与疫苗毒株通血清型同源,则动物生成的PRRSV 抗体水平远高于与疫苗毒株异源的攻击毒动物组。
Fu-Hsiang Hou(2020)等评估了同型异谱系的交叉保护力,采用Fostera株 PRRS MLV对谱系3分离物异源攻击进行交叉保护试验。免疫攻毒后检测了仔猪体温、呼吸道表现和病毒血症情况,结果见图2。

图2 各组在攻毒后体温、呼吸道表现和仔猪活力等的区别
从图2来看,在攻毒后,使用弱毒苗组(橙色)显著缩短了发热天数和发热程度,但与对照组相比,呼吸道症状依然明显。
从图3来看,虽然免疫组症状有缓解,但攻毒后病毒血症、病毒载量和未免疫组差别不大,并且很长时间带毒,推断用Foster株PRRS MLV对异源病毒攻击仅能提供有限的部分保护,说明该弱毒活疫苗对异源保护力不强。
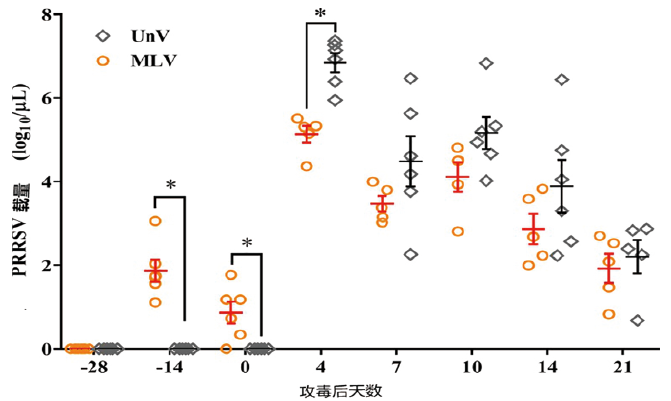
图3 血液病毒载量
Debin 等(2015)研究了不同嵌合毒株(一般认为其具有更好的交叉保护潜力)弱毒疫苗对异源攻毒的保护力,其评价有肺部组织病变和病毒载量的变化,结果见图4和图5。
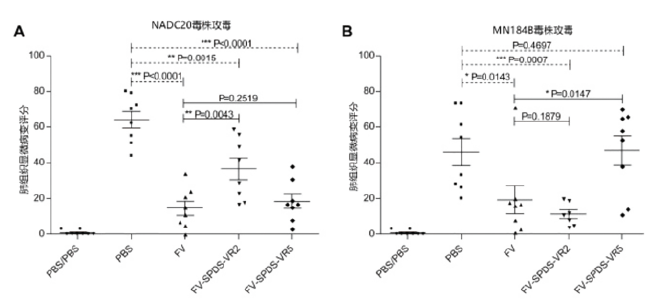
图4 肺部组织显微病变的评分
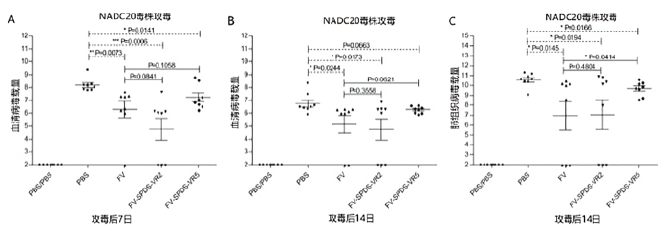
图5 肺部组织病毒载量
从结果来看,使用嵌合株弱毒疫苗,依然面临类似的问题,即不能很好地对异源毒株形成交叉保护,难以快速清除病毒,也无法快速的清除PRRSV。
在新型蓝耳弱毒疫苗的交叉保护力研究方面,Cindy(2019)等比较了普通活疫苗和含几种DNA免疫增强剂的活苗(FL13株)免疫仔猪,再通过鼻内接种异源毒株进行攻毒,采集血样和鼻拭子,检测病毒载量及血清抗体,以期待通过改进能增强活苗对异源病毒的保护力,结果见图6。
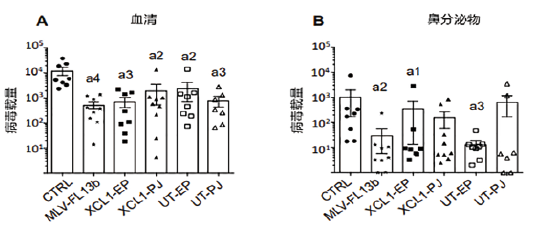
图6 血清和鼻分泌物病毒载量
笔者认为,与单纯MLV相比,DNA-MLV prime-boost方案在一定程度上改善了IFNγT细胞反应,并增加了针对包膜蛋白和N蛋白的抗体反应。但MLV和DNA-MLV免疫方案并没有进一步降低血清中异源病毒载量或鼻腔拭子的病毒载量。有研究表明,蓝耳1型和2型之间跨型的交叉保护力更是堪忧。
笔者注意到,很多文献发现有些弱毒疫苗对不同谱系之间毒株具有交叉保护力。交叉保护总会存在,只是强弱问题,但从总体上看,大多数猪蓝耳病活疫苗对异源毒株的保护力是有限的,对于大多数PRRS疫苗而言,其缺乏针对快速演变病毒的令人满意的异种交叉保护,这是明显的缺陷,所以选择PRRS疫苗时会感觉很难,而频繁更替PRRS活苗,出现的问题会更大。另一方面,有文献指出,遗传相似性没有固定的“截止值”来分类“同源”或“异源”毒株,PRRS MLV赋予同源保护的效力很难仅仅通过序列比较来预测,特别是仅基于GP5的序列,很多情况下ORF5的遗传同源性程度不是疫苗效力的良好预测因子。这是可以理解的,因为没有证据支持ORF5编码的GP5是唯一与保护相关的病毒蛋白,这也可以在一定程度上解释为什么部分猪场做了测序比对,然后选择相应弱毒苗免疫,控制效果依然达不到预期。
此外,鉴于PRRSV的广泛遗传和抗原变异,现场的大多数使用疫苗的情况都可被视为“异源挑战”,因为现场毒株与商业疫苗毒株或多或少不同。因此,在PRRS弱毒苗的合理使用方面依然要进一步探索。
当然,目前也有部分PRRS疫苗展示了较好的交叉免疫保护力,但是仍有很多问题值得我们去关注和反思,比如怎样才能正确有效的评价同源性以提供更好的免疫参考,如何避免养殖场测序比对选择疫苗但结果达不到预期等问题。
|
