|
不同年份2.3.4.4分支H5亚型禽流感病毒HA基因的序列分析摘要:H5亚型禽流感2.3.4.4分支病毒已流行了几年,当前的流行毒株是否会因变异而导致免疫失败引起关注。本研究对近年来流行毒株进行测序,比较分析了其抗原位点的变异情况,结果发现2017年的流行毒株已发生较大的变异,形成了一个独立的分支,与其之前的毒株HA核苷酸序列的差异达4.4%-5.4%。其最大特征是HA蛋白的142位E(按A/Goose/Guangdong/1/96毒株排序)发生缺失,导致该处增加了1个糖基化位点;同时受体结合位点的口袋右缘发生L145S的变异,而这一位点变异时间在2016年的毒株就已产生;2017的毒株另有12个aa发生了较为一致的变异。故推测当前毒株的抗原性已发生了较大变化。但238位(H3排序226位)匀为Q,保持禽源毒株的特征。需采取增加疫苗免疫频次及剂量,强化生物安全措施加以防控。
关键词:H5;2.3.4.4分支; HA;序列分析
H5亚型离流感自1996年发现以来已流行了20多年,给养禽业造成了巨大的损失,同时对公共卫生构成了严重威胁。病毒在这其间发生了多次的变异,产生了0-9分支,共10个不同分支的毒株。每间隔3-4年H5亚型流感病毒发生1次新的优势流行毒株的更叠,疫苗也随之不断更换。当前流行毒株主要为2.3.4.4分支,与其相对应的疫苗为Re-8疫苗,该分支毒株由2008年出现的2.3.4分支进化而来,2014年在韩国、越南等国流行以来, 已有几年,近期生产上也有免疫失败情况增多的反映,当前流行毒株是否发生了变异,引起特别关注。本研究选取日常监测中发现的阳性样测序结果进行分析,旨在了解当前毒株的变异情况,指导生产与防控工作。
1
材料与方法
1.1材料
1.1.1 样品:拭子样品8份,不同年份的采集于活禽交易市场。
1.1.2试剂:抽提试剂RNAiso Reagent、一步法RT-PCR扩增试剂(PrimeScript? One Step RT-PCR Kit ),购自宝生物工程(大连)有限公司。
1.1.3 HA基因的扩增引物:上游引物P1:5`-CAGGGAGCAAAAGCAGGGGTYCAAT-3`;下游引物P2:5`- GGTGGATTCTCTGTCTGCAGCGTAC-3`,P3: 5`-CTACTAGATCCCAAGTAAACGGGCAAC-3`;下游引物P4:5`-AGAAACAAGGGTGTTTTTAAYTACAAT-3`,上海立菲生物工程有限公司合成。
1.2方法
1.2.1 RNA的提取:按抽提试剂操作说明书进行。
1.2.2 HA基因扩增体系与程序:上、下游引物(25μM)各1μL、2×Bμffer 25μL、PrimeScript 1 Step Enzyme Mix 2μL 、DEPC水11μL和所提取的病毒RNA 10μL,体系共50μL,反应程序为50℃ 30min,95℃ 2min,然后95℃ 30s、53℃ 30s、72℃ 2min,30个循环,72℃延伸8min。
1.2.3序列测定和分析
将PCR产物送上海立菲生物工程有限公司进行测序,结果用DNAstar软件进行序列拼接。并与下载自GenBank 的禽流感病毒祖先代表株A/goose/donguan//2017/H5的HA基因组序列进行比较, 采用MEGA 5.0软件绘制其HA因组的系统进化树。
2
结果与分析
2.1HA 基因扩增结果
扩增出大小分别为1191bp和1057bp的特异性条带,与设计预期相符。见图1。
图1 PCR扩增结果
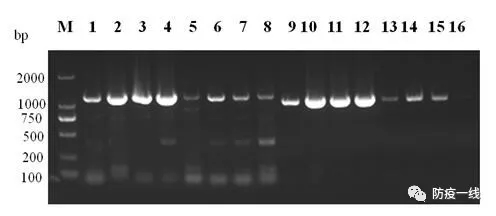
M、DNA Marker DL2000;1-8、P1P2扩增结果;9-16、P3P4扩增结果2.2HA基因遗传进化分析
将2017年的毒株A/goose/donguan//2017/H5,通过与Genbank上收录的序列进行比较,结果相似性最高的为KY415645(A/chicken/Ganzhou/GZ21/2015/H5N6),二者核苷酸的相似性为98%。从进化树可以看出近年毒株可分为2个分支。其中2017年的3个株毒已形成一个独立的分支;2013-2016年形成另一个分支,该分支各毒株间也有一定的差异,但不同年份毒株的差异不具有时间规律性。与最早出现的高致性H5代表株A/Goose/Guangdong/1/96(H5N1)相比有较大的差异,见图2。当前最新的2017年毒株与A/Goose/Guangdong/1/96相比,核苷酸差异已达10.1%-10.2%,与2016年毒株相比差异性为4.2%-4.8%,与2015年毒株相比差异性为5.1%-5.4%,与2014年之前的毒株相比差异性为4.4%-4.5%。2017年3个毒株间的差异较小,仅为0.4%-0.7%。2016年以前的毒株间的差异相对较小,2.1%-3.8%。
图2 H5HA基因的系统进化树
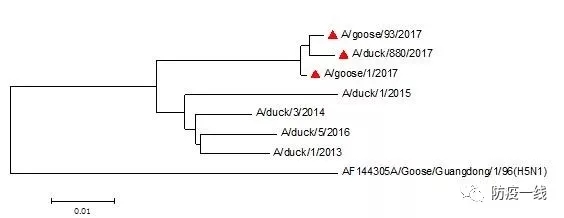
▲为2017年毒株
2.3HA 基因氨基酸序列分析
2017年病毒的cDNA 均包含了一个完整的开放阅读框架(ORF),编码区长1701,共编码566个氨基酸(aa),包括信号肽( 1-16aa) ,HA1 (17-343aa),HA2 (345-566aa)和1个精氨酸(344aa)。2016年以前的毒株则在HA1多1个氨基酸。
2.4糖基化位点与裂解位点的分析
氨基酸序列分析结果表明,2016年以前的毒株HA基因的潜在糖基化位点有7个,分别位于 26-28、27-29、39-41、 181-183 、302-304、500-502、559-600位aa位点上,位点1-5位于HA1,位点6-7位于HA2。2017年毒株潜在糖基化位点增加至8个,在140-142位增加了1个位点,主要是由于2017年毒株发生142位E缺失后产生,这是近期毒株的重要变异之一。裂解位点方面,各毒株呈现强毒的分子特征,PLRERRRK/RGLF,碱性氨基酸较A/Goose/Guangdong/1/96株少1个,详见表1。
表1 潜在糖基化位点分析
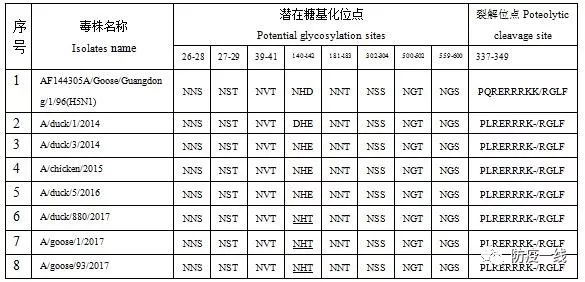
注:序号2-8为参考毒株。有下划线的为新增的位点。
2.5受体结合位点分析
HA蛋白的受体结合位点(Receptor Binding Site,RBS)是病毒感染识别和结合细胞受体的部位,HA三聚体的每1个单体都具有1个RBS,包括口袋左缘236-240位(NGQSG),口袋右缘145-149位(SGVSS)、口袋底部107位Y和165位W、口袋后壁202位E和206位L。本研究中2.3.4.4毒株的受体结合位点的较一致,但从2016年的毒株开始,在口右缘发生了L145S的变异,这可能导致宿主范围和抗原性发生改变。238位(H3排序226位)匀为Q,保持禽源毒株的特征。2017年以后的毒株另有12个位点发生了较一致的变异,主要集中在HA1肽段,其中142位E出现了缺失是一个重要的特征。56、131、139和145位较之前2.3.4.4分支的毒株发生了变异,但与H5亚型流感的祖先毒株A/Goose/Guangdong/1/96株一致,呈现出返祖的现象,推测其抗原性较早前的毒株更为接近。详见表2。
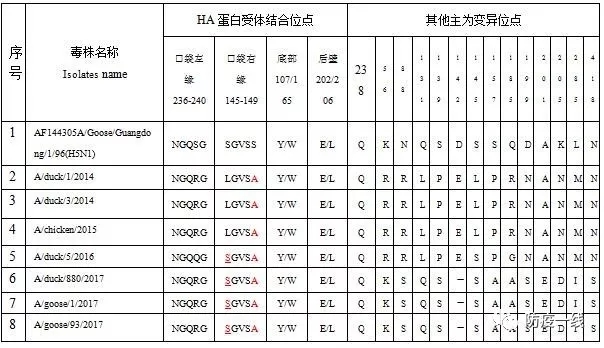
表2 HA受体结合位点及其他变异位点分析
注:1.HA蛋白氨基酸序列按A/Goose/Guangdong/1/96毒株;2.238-240位氨基酸分别对应H3排序的226-228位氨基酸;3.加下画线的为变异的位点。
3
讨论
3.1我国在2004年禽流感之后普遍采用疫苗免疫加扑杀作为主要的防控手段,在高强度的免疫压力下,增加了病毒变异的可能性,导致各种新流行株的出现。疫苗也随之不断更换,当前使用的Re-8已推出2年,部份地区有免疫失败的反映,当前流行毒株的变异情况是否达到导致免疫失败的程度,成为关注的焦点。本研究通过对HA基因核苷酸同源性分析发现2017年的流行毒株已较以往毒株已发生较大的变异,单独处于一个分支上,核苷酸序列较2016年以前的毒株差异性为4.2%-5.4%,与祖先毒株A/Goose/Guangdong/1/96相比,核苷酸差异已达10.1%-10.2%。2017年后出现这一较大的变异很可能导致与当前使用的疫苗株的匹配性较大幅度下降。
3.2 进一步对推导的HA蛋白的氨基酸序列进行分析发现2.3.4.4分支的流行毒株2017年后发生多个位点的变异。其最大特征是142位E(按A/Goose/Guangdong/1/96毒株排序)发生缺失,同时导致该处增加了1个糖基化位点140NHT142,使潜在糖基化位点增加至8个。受体结合位点中口袋右缘发生L145S的变异,而这一位点变异时间在2016年的毒株就已产生,2017的毒株另有12个位点发生了较为一致的变异。故推测当前毒株的抗原性已发生了较大变化。有趣的是2017年毒株在12个变异位点中有4个位点的呈现出返祖变异现象,故推测当前流行毒株的抗原性与较早前的流行毒株更为接近。
3.3 2014年在四川省出现首例人感染H5N6病例以来,在云南、广东、湖北和安徵引起多例人的感染,本研究中6个毒株HA蛋白238位(H3排序226位)匀为Q,保持禽源毒株的特征,人源毒株也有相似的情况。还与其他基因位点有关,不排除其他位点的改变引起宿主范围的变化。
3.4 关于防控措施。根据以往的流行规律,在流行毒更叠时期,水禽的风险较大,需引起特别关注。由于水禽是流感病毒的自然宿主,在长期的生物进化过程中对流感病毒的敏感性下降,疫苗免疫反应较差,导致产生的抗体水平不高,一旦流行毒株的抗原性与疫苗毒株的匹配性下降,水禽的发病风险则较大,由于疫苗毒株的更换需要一定的条件和时间,故当前只能加大免疫频次和剂量。水禽特别是樱桃谷鸭等,虽然饲养周期短,最少进行两次以上的免疫,剂量适当加大,例如7-10d 0.5mL,13-20d 1.2mL。同时做好生物安全措施,特别是隔离消毒好与市场有关的车辆与笼具。
作者:广东省动物疫病预防控制中心 卢受昇,孙彦伟,查云峰,叶健,吴立炀
|
