|
摘要
环境感染源可在形成流行病学动态方面发挥主要作用;然而,很少有人统计环境传播对宿主-病原体系统的相对影响。我们建立了一个非洲猪瘟病毒在野猪中的空间显式模型,并对其进行了拟合,估计东欧野猪中非洲猪瘟病毒的低水平持续存在,有多少比例是以尸体为基础的传播导致的。我们的模型是基于生态观察以及来自波兰东部非洲猪瘟和野猪的野外研究数据建立的。我们预测,基于尸体的传播将在疾病持久性方面发挥重要作用,特别是在接触率低的低密度宿主种群中。通过使用近似贝叶斯计算方法将模型与疫情数据进行拟合,我们推断出53%至66%的传播事件是基于尸体的,即通过宿主与受污染尸体的接触传播的。模型拟合和敏感性分析表明,随着宿主密度的降低,以尸体为基础的传播频率增加。这表明治理策略应强调尸体的清除,并考虑宿主密度的降低如何驱动以尸体为基础的传播。敏感性分析还表明,在真实参数下,基于尸体的传播对于非洲猪瘟的自主持久性是必要的。通过直接传播的自主持久性需要较高的宿主密度;否则,当直接传播概率中等高时,需要定期重新引入病毒才能持久。我们使用容易收集的生态数据和病毒监测数据,量化低流行疾病不同持久性机制的相对作用。了解不同传播机制的频率如何随宿主密度而变化,有助于在不断变化的生态条件下确定最佳治理策略。
关键字:非洲猪瘟,近似贝叶斯计算,尸体,环境传播,持续性,空间模型,传播,野猪
1
前言
了解病原体在宿主之间传播的机制是确定疾病风险和规划有效控制策略的关键。除了宿主与宿主直接接触或媒介传播外,病原体还可通过环境来传播,如接触污染物、饮用受污染的饮用水、接触受污染的土壤、接触受污染的尸体或清除尸体。增加环境传染源与易感宿主接触的可能性,可以促进病原体的持久性,因为许多病原体在环境中存活的时间比宿主感染的时间长。例如,流行病学模型表明,当传染源在环境中仍可存活时,病原体可以以很小的流行率持续存在于少数人群中。对于季节性密度变化的野生动物种群,易感宿主不足以通过直接接触持续保持病原体传播时,环境传染源可在低密度时期引发季节性流行病。感染源在环境-宿主体内的持续存在,使宿主的感染期超过宿主的预期寿命,从而在低流行增长率的情况下,使病原体的繁殖数量增加。在一些系统中,环境传播机制可以解释周期性流行,即使间隔时间长于种群周期,也可以通过增加群体之间的接触感染机会来增加群体间传染率。种群模型已经表明,除了直接传播的途径外,考虑环境传播的机制还可能导致疾病的动力学变化,并预测不同的动物运动阈值,从而导致种群减少。虽然确定最佳疾病管理策略需要运用传播机制的知识来确定适当的控制点,但对大多数宿主-病原体系统而言,量化环境传播的相对作用相较于其他传播机制是困难的。
非洲猪瘟(ASF)是一种高致病性的疾病,对养猪业和全球粮食安全造成严重影响。已知该病毒通过宿主间的接触、接触受感染的尸体、肉制品、污染物、气溶胶进行传播或通过蜱媒传播。尽管非洲猪瘟在非洲流行地区的森林宿主中持续处于低水平,但在没有家猪种群病毒扩散或其他感染源的情况下,非洲猪瘟在东欧野猪种群中持续低水平存在的状况仍令人费解。考虑到野猪经常接触同种动物的尸体,因此假定具传染性的尸体能够使非洲猪瘟在野猪种群中持续存在。以尸体为基础的传播是环境传播的一种特殊情况,其污染来自生物材料。这种传播被假设为一种低水平持久性的潜在机制,因为相对于活的感染宿主,尸体可以保持长时间的感染性。第二个假说解释了非洲猪瘟病毒在野猪种群中持续存在的原因,即来自邻国的持续输入对非洲猪瘟病毒的持续存在起了作用。为了评估这些假设,我们使用近似贝叶斯计算(ABC)开发了一个空间显式的机制流行病学模型,并将其与时空疾病监测数据(图1)拟合。我们估计了直接传播、基于尸体的传播和持续输入的水平,这些水平最能解释非洲猪瘟病毒的空间传播模式。我们对传播机制参数进行敏感性分析,以了解宿主密度与基于尸体传播之间的关系。我们假设,由于野猪一般不会移动太远,基于尸体的传播很可能占了整个传播的很大一部分。同样,我们预测,随着宿主密度的降低,尸体传播的潜在作用将增加,因为在低宿主密度下,由于野猪的短程运动趋势和病毒的感染期较短,直接接触将更加有限。
图1 波兰野猪群体中非洲猪瘟暴发的地点(2014-2015年)
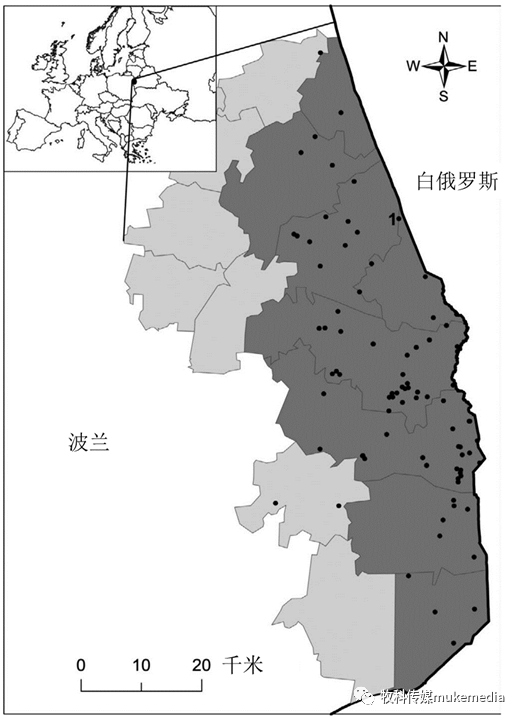
黑点表示野猪非洲猪瘟病例,并标有第一个病例。阴影区域表示用于参数估计的监测数据来源的行政区域(文本中的深灰色“感染区”,浅灰色“缓冲区”)
2
方法
2.1 波兰的非洲猪瘟
2014年2月在波兰东北部(北纬53°19′33",东经23°45′31"),距离与白俄罗斯边境不到1公里处发现了第一例非洲猪瘟野猪病例(图1)。在波兰首次出现非洲猪瘟之后,在受影响地区实施了强化监测策略。该策略是基于对所有死于交通事故的野猪(被动监测)和所有被猎杀的野猪(主动监测)的实验室检测。2014-2015年间,波兰共捕杀了4625头野猪,采集了271具尸体样本(图1)。由兽医服务机构和猎人采集的样本提交给位于波兰普阿维的国家兽医研究所的非洲猪瘟病毒诊断实验室。我们使用了2014年至2015年间在波兰8个行政区中非洲猪瘟病毒的监测数据(图1)。在此时间范围内,距离边境约100公里的区域中,共有2,761份野猪样本中,139份检测为非洲猪瘟病毒阳性(图1)。距离边境最远的是2015年底发生的第139起案件,距离边境34.5公里。由于抽样与狩猎活动和随机发现的尸体有严格的联系,监测数据是不定期和季节性变化的。关于实验室程序和测试的详细说明,请参见Woźniakowski等人,2016。
2.2 建模方法
为了解释野猪种群动态及其对非洲猪瘟病毒生态学的影响,我们建立了一个基于个体的空间显性模型,该模型考虑了个体在空间行为和社会互动中的差异。使用近似贝叶斯计算方法将模型与来自波兰的非洲猪瘟病毒监视数据进行拟合,从而估计未知参数(如下所述)。估计了新的病毒引入率和基于尸体的直接传播的概率。从白俄罗斯引进病毒的比率从每年1次到60次不等(即在边境不断蔓延)。同样,对于直接和基于尸体的传播机制,考虑了先验分布,其分布范围为每天从0%传输概率到100%的传输概率。在没有预先了解这个特定系统中的传输动力学的情况下,评估这三种持久性机制在解释观察到的监视数据方面的贡献。由于极端的计算要求,对一部分数据进行了敏感性分析,以广泛评估病毒维持机制在宿主种群密度范围内的变化。所有分析均在Matlab中进行。
2.3 个体模型
2.3.1 景观
景观由5×5km(25km 2 )的网格单元组成,网格单元的排列方式与疫区相似(图1,图S2)。网格单元的承载能力为0.5-2头公猪/km 2 ,通过密度依赖 性繁殖控制了整个环境的种群密度异质性。总景观面积为120×50km(6000km 2 )。
2.3.2 属性
每天都会对个体属性进行监测和更新。属性包括疾病状态、年龄、性别、独特的群体识别、散布年龄和距离、生命状态、繁殖、自然死亡年龄、x坐标、y坐标和网格细胞ID。模型中包括以下疾病状态以追踪非洲猪瘟病毒传播:易感、暴露、感染(存活)和感染(死亡尸体)(图2)。性别、扩散距离和自然死亡年龄在整个生命周期中是固定不变的,但其他属性根据时间、年龄、群体大小和网格密度而变化。空间在个体层面上是连续的。生命状态被监测为存活或死亡。生殖状态描述了女性基于年龄的受孕能力、妊娠状态和自上次分娩以来的时间。根据以下过程更新个体属性:自然死亡率、疾病传播、传播和社会动态、监测采样和繁殖。
图2 疾病状态转变和种群流动示意图
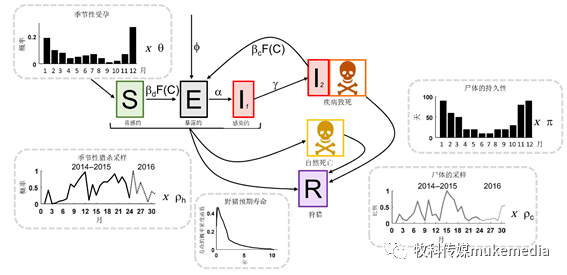
条形图显示了出生率和尸体持久性的季节性趋势。线图显示了狩猎和尸体采样强度的季节性趋势。共有三种死亡机制:疾病致死(I 2 ),自然死亡或狩猎。有两种潜在的传播途径:直接传播(d)或基于尸体的传播(c),它们是通过空间接触函数F(C)和给定接触的传播概率β(直接传播,β d 和基于尸体的传播,β c )发生的。尸体在环境中的持久性随季节而变化(反映了随天气而变化的降解率),但不论死亡机制如何,都是一样的,因此所有死亡机制的尸体都有相同的采样概率。受孕概率,尸体持久性和采样强度的季节性趋势都乘以估计的缩放参数(θ,π,ρ h ,ρ c )。我们还允许以频率ϕ在东部边界引入暴露个体。
2.3.3 疾病动力学
利用感染方程(FUI;易感个体感染率)来模拟疾病传播,其中x i ,j是传染性个体i(I i )的范围质心与易感性个体j(S j )(由其x和y坐标定义)之间的距离,a表示存活个体,b表示传染性尸体,d表示直接传输,c表示基于尸体的传输,β是特定于传输机制(d或c)的传输概率。为了说明野猪的空间接触行为,根据速率参数λ,假设传播概率随距离呈指数衰减(表1)。此外,野猪由于家族分组而呈现出异质的接触方式,因此,如果接触发生在同一家族(β wd 和β wc )内,则直接传播和基于尸体的传播的可能性更大。在等式1中,w表示在同一族组内的I i -S j 触点。具体参数见表1。
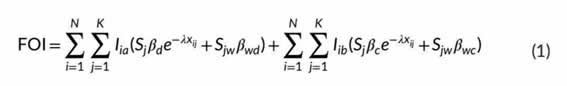
大量研究表明,猪科动物极易感染多种非洲猪瘟病毒,近100%的家猪和野猪在暴露后8-20天内死于疾病。病毒排出持续超过6周,感染也可能以慢性形式出现。但是,东欧地区从未报道过导致慢性感染的减毒病毒株。考虑到存活野猪宿主的生存能力在很大程度上是未知的,并且只有一小部分个体可能在病毒暴露下存活,我们假设感染非洲猪瘟的野猪中是100%致死的。宿主的潜伏期和感染期是随机变量(表1)。我们将病毒在尸体中的存留时间与尸体的腐烂率联系起来,以使未感染和受感染的尸体有相同的采样机会(我们找不到其他数据来证明这一点)。因此,我们认为被感染的尸体在环境中存活的整个期间都具有传染性。根据波兰东部尸体持久性的实地测量,假定尸体的感染期随季节而变化(表1,图2)。
表1 模型参数

2.3.4 社会结构与传播
野猪的社会结构是建立在母系群体基础上的,由少数亚成体和成年母猪及其后代组成。研究表明,内部个体之间的直接接触频率要比群体间的直接接触频率高得多。此外,社会群体可能会暂时分裂、改变或个体交换,但群体成员通常会形成稳定而持久的关系。因此, 社交和空间运动行为可以限制野猪的接触并调节传染病的传播。如图 3所示,我们考虑了社会结构的影响。雌性和未成年雌性在一个家庭群体中。同一家族的成员具有相同的质心。成年雄性是独立的(不是一组的一部分,每个人都有一个独特的家庭范围的质心)。社会结构是动态的家庭群体,组太大时(根据最大群体大小参数;表1)分为两组,一组分散(图S3)。同样地,因为成年母猪很少单独出现,所以单独的母猪被合并到能力不足的家庭群体中(表1)。从威布尔分布中随机选择分散距离(表1; 图S3)。公猪独立分开,母猪之间分散。尽管离开母猪的幼年野猪的散布是长距离迁徙的主要来源,但大多数个体在相对较短的距离(直径1-3公里)内(<5平方公里)散布,较长的散布(5-30公里)则不太常见。散布过程如下:(a)对于离初始质心的每个45度角,使用随机分配给散布点的散布距离值,获得新的[x,y]坐标集(表1;即x =距离x cos(角度)+当前x坐标,y =距离x sin(角度)+当前y坐标)。如果这些潜在地点中至少有一个是有效的(即,在一个公猪数量少于承载能力或远离网格的位置的网格单元中),然后随机选择一个有效的潜在位置,公猪(s)被重新安置在那里。离开网格的公猪永远地消失了。如果没有有效位置,则将距离值加倍,然后重复该过程,直到获得有效位置为止。
2.3.5 出生和死亡参数
根据季节性变化的受孕概率,生殖活跃的母猪随机发生受孕(表1;图2)。妊娠115天后,怀孕的雌性产下6个后代(3个雄性,3个雌性)(表1)。出生后,有3个月的固定滞后时间才有可能再次受孕(表1)。因此,每年最多产仔2窝。通过将受孕概率的季节性趋势乘以缩放参数(Ɵ),可以控制净人口增长率。先验分布Ɵ允许人口密度为承载力的10%时的净人口增长率在1.3到2.3之间。受孕概率与密度有关,因此受孕不会发生在已经处于承载能力的网格个体中。种群水平的宿主种群动态与logistic模型相似。
死亡率的来源包括自然死亡率,疾病引起的死亡率和猎人的狩猎(如下所述)。对于自然死亡率,每个个体在出生时根据野猪的预期寿命被赋予了一个寿命(表1;图2)。
2.3.6 初始条件和人口老化
种群初始化如下。创建了行数与所需总体大小相等的矩阵。每个个体(行)随机分配属性(表1)。对于年龄超过传播年龄的雄性,传播状态记录为完成。将所有小于分散年龄的雌性和雄性划分为组的1/4大小。每个个体或组被分配到一个随机选择的网格单元ID (该算法确保首先选择空闲的网格单元)。在每个网格单元中,个体或组被随机选择[x,y]坐标。初始化种群后,种群动态变化10年。以10年末的种群作为所有疾病传播模拟条件的起点。
2.3.7 监测参数
由于猎人的狩猎占抽样总数的大部分(94.5%),并且猎人的狩猎被认为是人口密度的主要调节因素,因此除了将其用作观察模型外,我们还将其作为死亡率的来源。2014年,估计野猪的平均密度为1.5-2.5只野猪/km 2 ,局部范围为0.5-1只野猪/km 2 至3-5只野猪/km 2 (国家森林区域管理局)。但是,由于没有关于公猪绝对数量与基础密度的关系的数据,因此添加了参数ρ h 和ρ c 来按比例缩放公猪的绝对数量(表1)。首先,计算了每种监测方法每天取样的公猪的相对数量(在t日取样的数量/任何一天的最大取样数量),以得出取样人口比例的季节性趋势(图2;图S4)。接下来,将每种监测方法的季节趋势数据乘以比例因子(ρh和ρc,表1;图S4),以确定通过猎人狩猎或从死亡尸体中取样的野猪的每日比例。趋势数据和比例因子的乘积可以视为每日检测概率。假设小于6个月龄的公猪不会被猎杀,小于3个月龄的公猪也不会被死尸法取样。记录了所有取样野猪的疾病状况,然后立即将它们永久性地从环境中清除。
2.4 近似贝叶斯算法
近似贝叶斯法计算使用距离度量(模型预测和观测数据之间的差异)为后验分布选择参数集,这是一种衡量模型参数集与观测数据中目标模式的近似程度的方法。我们同时使用三个距离度量;分别为: (a )来自尸体的每月病例数,(b )来自猎人采集样本的每月病例数,以及(c )每月距边境的最大距离的总和。距离度量公差值为:尸体的每月病例数为48,猎人采集样本的每月病例数为24,每月距边境的最大距离的为120。所有3个指标的结果低于这些值的参数集,构成后验分布。这使得平均错误率为2例(尸体)和1例(猎人狩猎),平均每月离边界5公里。根据我们认为可接受的误差水平来选择公差值,用于规划控制策略和风险评估。此外,更严格的错误率将需要大量的计算资源,除非先验分布得到更多的信息。
分别将模型拟合到4个不同的景观中:“斑片”包括高(2 公猪/km 2 )和低(0.5 公猪/km 2 )密度斑片,根据实际数据中病例的位置(平均密度~1公猪/ km2 )以及密度分别为1、1.5和2公猪/km 2 的“同质”景观。本设计评估了观察到的暴发模式是否是由于在观察到疾病的斑块中野猪密度高于其他斑块的潜在分布,而不是监测模式等替代机制造成的。
2.4.1 先验分布
每个参数具有如下均匀的先验分布:ϕ〜Unif(0,60),β d 〜Unif(0.0001,1),β c 〜Unif(0.0001,0.99),ρ h 〜Unif(0.0005,0.1),ρ c 〜Unif(0.0005,0.8),π〜Unif(0.1,1.5),θ〜Unif(0.5,6),λ〜Unif(0.1、2.5),β wd 〜Unif(0.01,1)和β wc 〜Unif(0.001,1)。先验分布范围由运动和接触数据决定。作为参数生成过程的一部分,我们定义β d >β c ,β wd >β d ,β wc >β c ;进一步利用生物知识说明先验分布。为了有效地跨参数空间进行采样,我们使用了拉丁超立方体算法生成979592个参数集,然后在每个参数集上对模型运行两次(共计1959184个迭代;或2个链的979592)。对β d 、β c 和ρ c 按log e 进行抽样。由于流行病学模型是时间密集型的,使用两层方法来评估参数集。首先,如果轨迹不符合实际情况,模拟提前终止。具体标准如下:(a)整个景观范围内的宿主密度下降到初始密度的20%以下,(b)每天发生150多个新病例,(c)过去6个月内没有新病例通过任何一种监测方法取样,或(d)两种监测方法抽样病例共300余例(比实际数量增加一倍以上)。然后我们只考虑模拟达到两年时间框架结束的参数集,对于减少的参数集,后验分布由所有唯一参数集(包括两个链)组成,这些参数集均在三个度量的绝对距离之内:来自活体和死亡动物的每月阳性样本(单独考虑)的观察数据与模拟数据之间的绝对差之和,以及病例离边界的每月最大距离。
2.4.2 拟合优度检验
为了确定“最佳”景观模型,根据不同景观的距离度量和R 2 值(观察到的与预测的每月病例和每月到边界的距离)对模型进行了排序(表1 )。为了计算R 2 值,从参数的后验分布中随机抽取1000个景观进行模拟,并计算每个模拟的R 2 值。然后,将每个指标的1,000个R 2 值的平均值(每月的情况和距离)表示特定模型的指标的总体R 2 值。作为预测能力的另一个衡量指标,使用2014-2015年数据拟合度估算的参数预测2016年的前7个月(1月至7月),测试模型预测非洲猪瘟动态的能力。然后,将样本内预测的R 2 值相对于整个预测集进行了比较(表1)。
2.5 灵敏度分析
对密度为1-4头野猪/km 2 的同质景观进行敏感性分析,反映了东欧野猪的观测密度。完成了完整的阶乘敏感性分析,以评估非洲猪瘟持续性和传播动力学对ϕ,βd和βc的变化的响应。传播参数β d 和β c 从无传播(0.0001)到高传播水平(0.3)不等,ϕ从每年1次引入到50次引入变化。所有其他参数均由斑块景观模型的后验分布参数集确定。利用3个不同的50km×50km景观(1-4头公猪/ km2 )进行敏感性分析。索引案例发生在网格单元50(网格单元最右侧列的中间)的第30天(在ABC分析中引入的同一天)。所有的实验都进行了2年。为每组进行了100次重复模拟。,并对所有情况进行记录。,包括因狩猎而导致的宿主死亡以及由于监测采样而导致的死尸清除。并记录发生以下情况的数据: (a )持久性概率(在100个模拟中,在开始时只有1次引入之后到两年时间内的最后一周至少出现1个案例的比例),以及(b )来自直接传播和基于尸体传播的传播事件的比例。后者的输出是通过记录每天直接传播的传播事件的比例,并取时间的中位数(仅考虑发生至少一个传播事件的天数)而获得的。使用广义线性模型对输出建模,对四个响应变量的每个变量使用适当的分布和/或数据转换,包括将传输概率参数和引入频率作为协变量,以及所有的交互作用。这些模型的目的仅仅是在模拟中的数值范围内达到更高的插值分辨率关系。对于持久性概率的建模,还包括了高达4阶的交互作用,因为这些关系是高度非线性的,因此这些对于精确地插值关系非常重要。
3
结果
3.1 模型拟合
尽管在几个估计的参数中存在很高的不确定性,但是使用这些模型可以很好地观察监视数据的总体趋势(图3)。所有模型每月捕获的病例都比每月距边界的最大距离要好(表2,图3)。相对于观察数据,该模型预测14-16个月(包括出生脉搏的第二年2-4月)的发病率较高,而第5-7个月(包括异常低监测期的第一年5-7月)的发病率较低(图3a,图S4)。模型中通过猎人狩猎和尸体采样,监测样本的真实患病率,监测观察到的平均患病率(图S5)。宿主密度均匀的景观模型没有采集到空间扩展率,也没有采集到高密度斑块(2 公猪/km 2 ; 平均1野猪/km 2 ; 表2 )。四种模型的参数集的拒绝率都很高,后验分布在6-53个值之间(0.00031%-0.0027% 的模型接受率)(表2 ),参数估计的不确定性很大(见图S6 )。由于模型处理的随机性和参数估计的不确定性,模型平均拟合数据(即相对于观测数据的随机运行轨迹中值R 2 ;图3c和d )优于任何一条轨迹的观测数据(即,每次随机运行的中值R 2 ; 图3a和b)。完整数据(包括样本外预测)的R 2 低于样本内预测(表2,图3a和b),表明模型在样本外的预测时表现更差。后验分布揭示了参数的相关性(图S7)。β d 和β c 彼此负相关,与λ负相关,而ρ c 和π正相关 (图S7)。其他参数相对不相关。
3.2 尸体传播的作用
这些模型预测了大量的基于屠体的传播(2014-2015年期间的月平均值在53%到66%之间,具体取决于景观;图4),采样尸体中的非洲猪瘟患病率远高于猎人狩猎采集的样本 (图S5a)。斑块状景观也预测了野猪种群随时间的缓慢下降 (图S5b),这反映,随着时间的推移,源自尸体传播的事件更多(图4),特别是在斑块状和低密度的均匀景观中。
图3、模型拟合(斑驳景观)
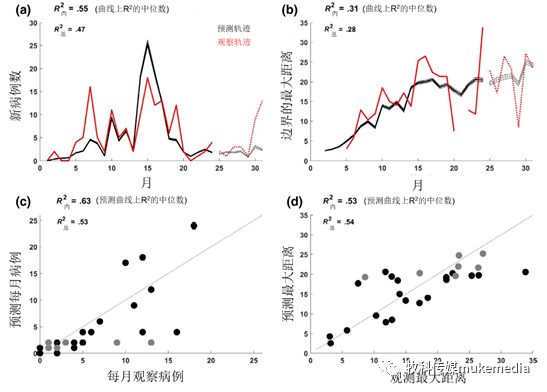
新病例的轨迹(a)和观察(红色)和预测(黑色)到边界的最大距离(b)。阴影区域表示根据参数的后验分布从1,000次模拟中得出的95%可信区间。实线表示用于参数估计的数据,而虚线表示样本外的预测。c和d显示了每月情况(c)和距边界的最大距离(d)的观察点与预测点(其中每个点是每个时间所有模拟的中位数)。样本内预测点(2014–2015)为黑色,样本外预测点(2016)为灰色。灰色虚线表示点的预期拟合度(观察点和预测点的比例为1:1)
3.3 宿主密度对非洲猪瘟持续性的影响
敏感性分析表明,密度高于1头公猪/km 2 对疾病持久性很重要(图5a、d和g)。如果没有基于尸体的传播,在较低的宿主密度下,疾病持久性需要每年重新引入10次或更多次(图5b和e)。然而,只有基于尸体的传播,即使在很少或没有重新引入时,在低宿主密度的情况下(图5c和f),非洲猪瘟也会在狭窄的基于尸体的传播概率范围内持续发生。相比之下,高宿主密度(4头公猪/km 2 )即使在没有尸体传播的情况下(图5g-i),在任何一种传播机制的狭窄的传播概率范围内,可以实现疾病的持续性存在。
表2、野猪种群不同密度和斑块度的模型选择和拟合
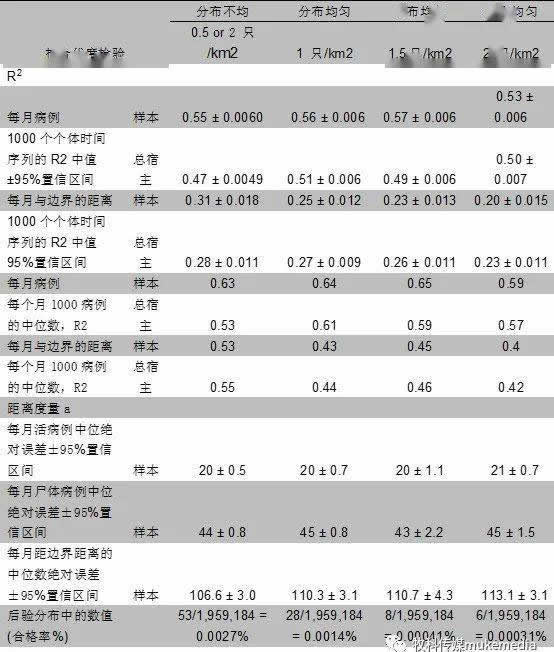
最能解释数据的模型以浅灰色突出显示。与其他模型相比,该模型产生了与案例数据相似的拟合,但在拟合到边界距离的R2和ABC距离度量方面表现明显更好
a:1000个模拟的后验分布的中位数距离度量值±95%置信区间。
图4、来自直接传播的传播事件的比例

阴影线是来自每个模型后验分布的1000次模拟的95%预测区间。红色表示>50%的传输事件是基于尸体传播;蓝色表示>的50%是直接传播。不同的线条显示了不同景观的结果。注意,参数估计值随地形的不同而不同(表2)。透明阴影面板表示样本外预测。
图5、宿主密度和持久性机制的影响
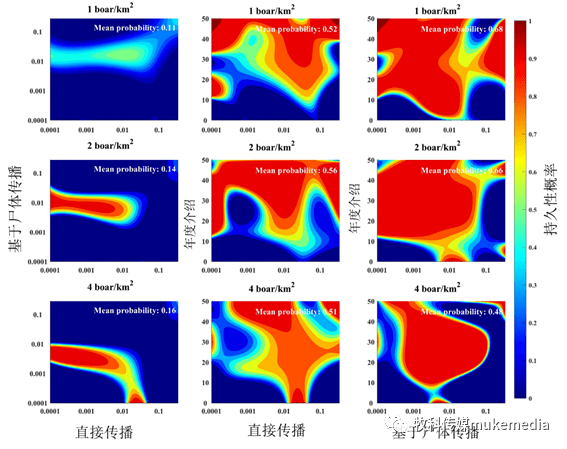
颜色显示ASFV持续存在的概率,深红色代表高概率,黄色代表中度高概率,浅蓝色代表中度概率,深蓝色代表低概率。
β d :群体直接传输概率,β c :组间基于尸体的传输概率,ϕ:引入频率。组内传播概率固定为组间传播概率的10倍。模型估计的其他参数固定为生物学上的实际值:ρh= 0.015,ρ c = 0.025 ,π= 1,θ= 2,λ= 1.5;其他参数如表1所示。每个图显示不同宿主密度的结果。黑色的平均值表示整个图的平均值,给出了景观对空间分布的整体影响。
4
讨论
东欧野猪中非洲猪瘟病毒的持续存在仍然对全球家猪种群构成重大威胁,因此也威胁着国际贸易和粮食安全。利用野猪种群动态的先验知识,将机械性疾病动态模型拟合到时空疾病监测数据中,我们推断53%-66%的非洲猪瘟病毒传播事件是通过易感宿主与死亡尸体的接触而发生的。在尸体腐烂的后期,野猪也会继续与尸体接触,因此加强对尸体的监测和清除可能会大大减少传播。因此,开发经济有效的尸体检测和寻找方法可能是降低野猪种群传播率的关键。此外,由于我们发现传播机制的相对重要性主要取决于宿主密度,因此我们的结果强调了将野猪种群动态纳入考虑的重要性。尽管我们的模型可以很好地采集数据,但当模型尽可能地拟合关联度时,它低估了空间传播的速度。当它很好地符合空间传播速度时,它高估了发病率。因此,我们模型的结构缺乏非洲猪瘟病毒在野猪种群中传播的重要未知过程。 由于我们的建模框架考虑了空间接触和传播距离,我们很难同时拟合发病率和距离轨迹,这可能是由于长距离运动经常发生,足以在日常家庭范围或传播运动之外播散感染。这些结果并不令人惊讶,因为人介导的非洲猪瘟病毒传播在该地区继续发挥着重要的流行病学作用。一个可能的来源是猎人在处理传染性尸体时可能污染狩猎设备,然后在另一个地点引入传染性污染物。 实施加强的狩猎生物安全政策可能有助于减少猎人造成的非洲猪瘟病毒的空间传播。第二种可能是其他与传染性尸体接触的物种,例如,食腐的食肉动物、鸟类和苍蝇,并随后散布受污染的组织。尽管机械载体在非洲猪瘟病毒流行病学中的作用尚不清楚,但已知机械载体能促进许多疾病的传播,提示这一机制值得进一步研究。
正如引入完全易感人群的新疾病所期望的那样,我们的模型倾向于在引入指示病例后立即预测相对较高的发病率。然而,狩猎在最初的3-4个月内受到限制,这导致了在此期间观察到的病例数量较少。一旦狩猎禁令被取消,观察到的病例数量就会增加,潜在的动态更加一致(数据未显示)。我们使用总体样本量的数据说明了狩猎的时间趋势,但我们没有阴性监测数据的精确位置。所有样本的位置对于数据拟合非常重要,监视系统限制了我们对基本过程的观察。如果在现实中,监视采样位置在距离边界较近或较远的空间簇中移动,而不是所有样本随机分布,则有可能检测到距离边界较远的病例。空间扩散的真实速度快于我们模型中的预测观测速度,这表明考虑所有监视样本的空间位置可以帮助改善对空间扩散过程的推断,并有可能使我们了解传播机制。我们的分析强调了在观测过程中充分记录阴性样本原始数据和适当考虑完整抽样设计的重要性。
几项研究已经观察到,去除患病宿主的种群,目标病原体的患病率有所下降。但是,扑杀可能会产生其他后果,例如,病毒适应了低密度条件,或者长距离宿主运动增加,导致疾病扩散增加。我们的敏感性分析还表明,降低宿主密度可能会产生意想不到的后果。虽然宿主密度高可以通过直接传播实现疾病自主持久性,但基于尸体的传播可以通过有效延长寄主接触受污染尸体的机会,在寄主密度降低时实现持久性。因此,在制定扑杀计划之前,深入了解宿主-病原体传播生态学的影响是很重要的。
最近的一项研究发现,对于野猪(一种聚集在家庭群体中的社会物种)来说,空间上的清除家庭群体的所有成员,比随机扑杀个体更有效。虽然这个模拟模型没有考虑到由于扑杀而引起的运动响应,但它确实包括了疾病传播中社会结构的影响。由于密集狩猎会导致逃逸运动和群体间的混合,所以扑杀计划应考虑清除群体中的所有个体,以减少由于社会结构破坏而增加的远距离移动的机会。
我们假设非洲猪瘟在野猪中具有100%的致死性。事实上,该地区的监测数据表明,在暴发地区,有低于1%的幸存者检测出抗非洲猪瘟病毒抗体阳性(国家兽医研究所未公布的数据)。因为我们假设了绝对致死率,所以我们的模型预测,随着时间的推移,持续传播的非洲猪瘟病毒会导致宿主密度降低和基于尸体的传播水平升高。但是,如果该地区出现了致死性降低的非洲猪瘟病毒毒株,那么可能会出现不同的流行病学动态。具体来说,如果有更多的宿主存活,它们将可以繁殖,保持较高的宿主密度,仅通过直接传播就可能导致持久性或复发性流行。因此,对致死率变化的监视对于随时间推移,优化局部区域的控制策略很重要。
我们对野猪空间过程的实现是对现实的简化。我们假设个体每天都有可能在各个方向上与其他个体接触,接触概率随距离衰减。然而,在现实中,野猪的迁徙偏向于栖息地特征和相关个体。尽管这些迁徙会导致实际距离迁徙概率关系随时间变化规律与我们假设的不一致,但是迁徙到特定位置(例如,偏向特定资源或特定物体)会更重要,而不仅仅是整个距离-变化模式。事实上,最近的研究表明,对麋鹿的运动进行机械性统计可以对布氏杆菌病随季节和气候变化的时空流行情况进行预测。基于景观异质性的机械运动模型也已用于预测接触率最高的区域,这是疾病传播风险的另一项指标。适应性预测疾病传播风险在何时何地达到最高,对于以经济高效的方式在空间和时间上确定缓解策略的优先级至关重要。此外,虽然理论预测基于社会结构知识实施反应控制,有助于确定控制的有效性,但很少有人能先验地了解个体的层次关系状态。 了解迁徙运动、景观异质性和疾病传播之间的联系,有助于为优先监测和干预提供更实际(基于景观的)指导。
来源:牧科传媒
|
